| The GLIMMIX Procedure |
Example 38.16 Diallel Experiment with Multimember Random Effects
Cockerham and Weir (1977) apply variance component models in the analysis of reciprocal crosses. In these experiments it is of interest to separate genetically determined variation from variation determined by parentage. We analyze here the data for the diallel experiment in Cockerham and Weir (1977, Appendix C). A diallel is a mating design that consists of all possible crosses of a set of parental lines. It includes reciprocal crossings, but not self-crossings.
The basic model for a cross is 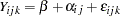 , where
, where 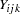 is the observation for offspring
is the observation for offspring 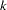 from maternal parent
from maternal parent  and paternal parent
and paternal parent 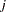 . The various models in Cockerham and Weir (1977) are different decompositions of the term
. The various models in Cockerham and Weir (1977) are different decompositions of the term  , the total effect that is due to the parents. Their "bio model" (model (c)) decomposes into
, the total effect that is due to the parents. Their "bio model" (model (c)) decomposes into
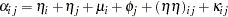 |
where 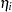 and
and 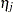 are contributions of the female and male parents, respectively. The term
are contributions of the female and male parents, respectively. The term 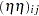 captures the interaction between maternal and paternal effects. In contrast to usual interaction effects, this term must obey a symmetry because of the reciprocals:
captures the interaction between maternal and paternal effects. In contrast to usual interaction effects, this term must obey a symmetry because of the reciprocals: 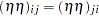 . The terms
. The terms 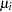 and
and  in the decomposition are extranuclear maternal and paternal effects, and the remaining interactions are captured by the
in the decomposition are extranuclear maternal and paternal effects, and the remaining interactions are captured by the 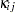 term.
term.
The following DATA step creates a SAS data set for the diallel example in Appendix C of Cockerham and Weir (1977):
data diallel;
label time = 'Flowering time in days';
do p = 1 to 8;
do m = 1 to 8;
if (m ne p) then do;
sym = trim(left(min(m,p))) || ',' || trim(left(max(m,p)));
do block = 1 to 2;
input time @@;
output;
end;
end;
end;
end;
datalines;
14.4 16.2 27.2 30.8 17.2 27.0 18.3 20.2 16.2 16.8 18.6 14.4 16.4 16.0
15.4 16.5 14.8 14.6 18.6 18.6 15.2 15.3 17.0 15.2 14.4 14.8 10.8 13.2
31.8 30.4 21.0 23.0 24.6 25.4 19.2 20.0 29.8 28.4 12.8 14.2 13.0 14.4
16.2 17.8 11.4 13.0 16.8 16.3 12.4 14.2 16.8 14.8 12.6 12.2 9.6 11.2
14.6 18.8 12.2 13.6 15.2 15.4 15.2 13.8 18.0 16.0 10.4 12.2 13.4 20.0
20.2 23.4 14.2 14.0 18.6 14.8 22.2 17.0 14.3 17.3 9.0 10.2 11.8 12.8
14.0 16.6 12.2 9.2 13.6 16.2 13.8 14.4 15.6 15.6 15.6 11.0 13.0 9.8
15.2 17.2 10.0 11.6 17.0 18.2 20.8 20.8 20.0 17.4 17.0 12.6 13.0 9.8
;
The observations represent mean flowering times of Nicotiana rustica (Aztec tobacco) from crosses of inbred varieties grown in two blocks. The variables p and m identify the eight paternal and maternal lines, respectively. The variable sym is used to model the interaction between the parents, subject to the symmetry condition . For example, the first two observations, 14.4 and 16.2 days, represent the observations from blocks 1 and 2 where paternal line 1 was crossed with maternal line 2.
The following PROC GLIMMIX statements fit the "bio model" in Cockerham and Weir (1977):
proc glimmix data=diallel outdesign(z)=zmat;
class block sym p m;
effect line = mm(p m);
model time = block;
random line sym p m p*m;
run;
The EFFECT statement defines the nuclear parental contributions as a multimember effect based on the CLASS variables p and m. Each observation has two nonzero entries in the design matrix for the effect that identifies the paternal and maternal lines. The terms in the RANDOM statement model the variance components as follows: line 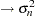 , sym
, sym 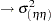 , p
, p 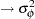 , m
, m 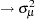 , p*m
, p*m 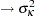 . The OUTDESIGN= option in the PROC GLIMMIX statement writes the
. The OUTDESIGN= option in the PROC GLIMMIX statement writes the 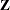 matrix to the SAS data set zmat. The EFFECT statement alleviates the need for complex coding, as in Section 2.3 of Saxton (2004).
matrix to the SAS data set zmat. The EFFECT statement alleviates the need for complex coding, as in Section 2.3 of Saxton (2004).
Output 38.16.1 displays the "Class Level Information" table of the diallel model. Because the interaction terms are symmetric, there are only 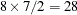 levels for the 8 lines. The estimates of the variance components and the residual variance in Output 38.16.1 agree with the results in Table 7 of Cockerham and Weir (1977).
levels for the 8 lines. The estimates of the variance components and the residual variance in Output 38.16.1 agree with the results in Table 7 of Cockerham and Weir (1977).
| Class Level Information | ||
|---|---|---|
| Class | Levels | Values |
| block | 2 | 1 2 |
| sym | 28 | 1,2 1,3 1,4 1,5 1,6 1,7 1,8 2,3 2,4 2,5 2,6 2,7 2,8 3,4 3,5 3,6 3,7 3,8 4,5 4,6 4,7 4,8 5,6 5,7 5,8 6,7 6,8 7,8 |
| p | 8 | 1 2 3 4 5 6 7 8 |
| m | 8 | 1 2 3 4 5 6 7 8 |
The following statements print the matrix columns that correspond to the multimember line effect for the first 10 observations in block 1 (Output 38.16.2). For each observation there are two nonzero entries, and their column index corresponds to the index of the paternal and maternal line.
proc print data=zmat(where=(block=1) obs=10);
var p m time _z1-_z8;
run;
| Obs | p | m | time | _Z1 | _Z2 | _Z3 | _Z4 | _Z5 | _Z6 | _Z7 | _Z8 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 2 | 14.4 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 3 | 1 | 3 | 27.2 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 5 | 1 | 4 | 17.2 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 7 | 1 | 5 | 18.3 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 9 | 1 | 6 | 16.2 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| 11 | 1 | 7 | 18.6 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| 13 | 1 | 8 | 16.4 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 15 | 2 | 1 | 15.4 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 17 | 2 | 3 | 14.8 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| 19 | 2 | 4 | 18.6 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
Copyright © 2009 by SAS Institute Inc., Cary, NC, USA. All rights reserved.