| The FAMILY Procedure |
| Statistical Computations |
For all tests, it is assumed that the marker has two alleles, 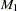 and
and 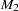 . Extensions to multiallelic markers are made by performing the tests on each allele in turn, with the current allele being considered to be and all other alleles considered to be . When the CONTCORR option is specified in the PROC FAMILY statement, the
. Extensions to multiallelic markers are made by performing the tests on each allele in turn, with the current allele being considered to be and all other alleles considered to be . When the CONTCORR option is specified in the PROC FAMILY statement, the  score statistics of all versions of the TDT, S-TDT, and RC-TDT can be continuity corrected by subtracting 0.5 from the absolute value of the numerator. The two-sided
score statistics of all versions of the TDT, S-TDT, and RC-TDT can be continuity corrected by subtracting 0.5 from the absolute value of the numerator. The two-sided  -value for each score using the normal distribution is equivalent to using the -value from the
-value for each score using the normal distribution is equivalent to using the -value from the 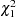 distribution for the square of the score, and this chi-square form of the statistic is reported in the output data set.
distribution for the square of the score, and this chi-square form of the statistic is reported in the output data set.
TDT
The TDT (Spielman, McGinnis, and Ewens 1993) is implemented using a normal approximation. This test includes families where both parents have been genotyped for the marker and at least one is heterozygous. If only one parent has been genotyped, that parent is heterozygous, and the affected child is not homozygous and does not have the same genotype as the typed parent, then the TDT can be applied to this family as well (Curtis and Sham 1995). The TDT tests for equality between the proportion of times a heterozygous parent transmits the allele to an affected child and the proportion of times a heterozygous parent transmits the allele to an affected child. The normal approximation to the binomial is used to form the score statistic
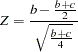 |
where 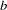 is the number of alleles in affected children from heterozygous parents and
is the number of alleles in affected children from heterozygous parents and  is the number of alleles in affected children from heterozygous parents.
is the number of alleles in affected children from heterozygous parents.
Two extensions to a multiallelic TDT are available. The first, which is performed by default or when MULT=JOINT is specified in the PROC FAMILY statement, combines the TDT for each of 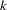 alleles at a marker into one statistic as follows (Spielman and Ewens 1996):
alleles at a marker into one statistic as follows (Spielman and Ewens 1996):
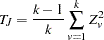 |
where 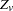 is simply the
is simply the  defined in the preceding paragraph, with allele
defined in the preceding paragraph, with allele 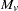 treated as and all other alleles as for each
treated as and all other alleles as for each 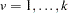 .
. 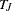 and the continuity-corrected form
and the continuity-corrected form 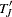 have an asymptotic
have an asymptotic 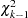 distribution, and the corresponding -value is reported.
distribution, and the corresponding -value is reported.
Alternatively, if the MULT=MAX option is specified, either 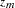 or
or 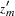 (when the CONTCORR option is specified) is used, where
(when the CONTCORR option is specified) is used, where 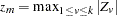 . The equivalent one degree of freedom chi-square statistic is reported, and a Bonferroni correction is applied to its -value.
. The equivalent one degree of freedom chi-square statistic is reported, and a Bonferroni correction is applied to its -value.
Note:The TDT is a valid test of linkage and association only when the data consist of unrelated nuclear families and each family contains only one affected child. Otherwise, it is a valid test of linkage only.
S-TDT
The score procedure given by Spielman and Ewens (1998) is used to calculate -values for the S-TDT. This test can be applied to families where there are at least one affected sibling and one unaffected sibling, and not all siblings have the same genotype. The score, whose two-sided -value is approximated using the normal distribution, is calculated as 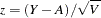 .
.  represents the total observed number of alleles in the affected siblings. For
represents the total observed number of alleles in the affected siblings. For  total siblings in the family,
total siblings in the family,  affected and
affected and  unaffected, and
unaffected, and  that are
that are 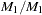 and
and  that are
that are 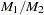 , summing over families gives
, summing over families gives
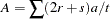 |
and
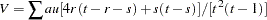 |
as the expected value and variance of , respectively.
When the COMBINE option is specified in the PROC FAMILY statement, the S-TDT and TDT are combined as follows: the TDT is applied to all alleles within a family that meet the requirements described in the preceding section. The S-TDT is then applied to the remaining alleles within a family that meet its requirements described in the preceding paragraph. Using the notation already given for these tests, the score for the combined test can then be written as
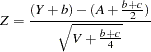 |
For multiallelic markers, the same extensions can be made to the S-TDT and combined S-TDT that were made to the TDT (Monks, Kaplan, and Weir 1998); that is, either a joint test over all alleles (using 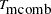 ) or the maximum score of all the alleles with the -value being Bonferroni-corrected.
) or the maximum score of all the alleles with the -value being Bonferroni-corrected.
Note:The S-TDT is a valid test of linkage and association only when the data consist of unrelated nuclear families and each family contains only one affected and one unaffected sibling. Otherwise, it is a valid test of linkage only.
SDT
The SDT (Horvath and Laird 1998) is a sign test used on discordant sibling pairs. As with the S-TDT, one affected sibling and one unaffected sibling are required to be in each family, but unlike the S-TDT, the SDT remains a valid test of linkage and association when the sibship is larger.
The notation from the S-TDT is used, except now the quantities 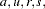 and are defined for each sibship/family, so, for example, there are affected siblings in the family and unaffected siblings in the family. Treating each allele in turn as and all other alleles as , , define for each family in the data the average number of
and are defined for each sibship/family, so, for example, there are affected siblings in the family and unaffected siblings in the family. Treating each allele in turn as and all other alleles as , , define for each family in the data the average number of  alleles among affected siblings and unaffected siblings respectively as
alleles among affected siblings and unaffected siblings respectively as
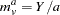 |
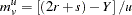 |
Then 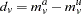 for each family, and summing over families gives
for each family, and summing over families gives 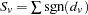 , where sgn
, where sgn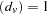 for
for 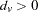 , 0 for
, 0 for 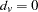 , and
, and 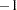 for
for 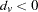 . The joint multiallelic SDT statistic (mSDT) is then defined by Czika and Berry (2002) as
. The joint multiallelic SDT statistic (mSDT) is then defined by Czika and Berry (2002) as 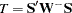 , where
, where 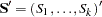 and
and 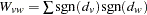 ,
, 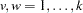 , and
, and 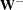 is the Moore-Penrose generalized inverse of
is the Moore-Penrose generalized inverse of 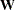 .
.  has an asymptotic
has an asymptotic 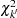 distribution, where
distribution, where  , and this distribution is used to obtain -values for the SDT (Czika and Berry 2002). When there are only two alleles at the marker, this joint multiallelic version of the SDT reduces to the biallelic version of the SDT.
, and this distribution is used to obtain -values for the SDT (Czika and Berry 2002). When there are only two alleles at the marker, this joint multiallelic version of the SDT reduces to the biallelic version of the SDT.
This sibship test is also combined with the TDT when the COMBINE option in the PROC FAMILY statement is specified, creating a test that can potentially use more of the data (Horvath and Laird 1998; Curtis, Miller, and Sham 1999). In order to maintain the test’s validity as a test of association in families with more than one affected and one unaffected sibling, a nonparametric multiallelic TDT is used, which is in the same 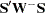 form as the SDT. This test statistic for the joint test also has an asymptotic distribution (Czika and Berry 2002), and the corresponding -value is reported.
form as the SDT. This test statistic for the joint test also has an asymptotic distribution (Czika and Berry 2002), and the corresponding -value is reported.
When the MULT=MAX option is specified in the PROC FAMILY statement, then the SDT chi-square statistic is simply 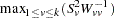 and has one degree of freedom. This applies to the SDT when used alone or combined with the TDT. As with the other tests, a Bonferroni correction is made to the -value.
and has one degree of freedom. This applies to the SDT when used alone or combined with the TDT. As with the other tests, a Bonferroni correction is made to the -value.
RC-TDT
The RC-TDT (Knapp 1999a) takes the combined S-TDT a step further by reconstructing missing parental genotypes when possible in order to use more families. The RC-TDT can be applied to families with at least one affected child that meet one of the following conditions:
Both parents are typed with at least one heterozygous for
. One parent is typed, the other can be reconstructed, and at least one parent is heterozygous for
. Both parents’ genotypes are missing but can be reconstructed, and at least one parent is heterozygous for
. At least one parental genotype is missing and cannot be reconstructed, but the conditions for the S-TDT are met.
One parental genotype is missing and cannot be reconstructed, the other parent is heterozygous for
, and at least one affected child is heterozygous for and an allele not in the typed parent (Knapp 1999b).
Reconstruction of parental genotypes is attempted only when there are no genotyping errors in the family for the marker being tested. As with the S-TDT, a score is created using the statistic , but Knapp (1999a) calculates a different expected value  and variance of , which takes into account the bias created by the genotype reconstruction, to form the score over all families:
and variance of , which takes into account the bias created by the genotype reconstruction, to form the score over all families:
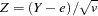 |
For multiallelic markers, the same extensions can be made to the RC-TDT that were made to the TDT and S-TDT–that is, either a joint test over all alleles, or the maximum score of all the alleles with the -value being Bonferroni-corrected.
Note:The RC-TDT is a valid test of linkage and association only when the data consist of unrelated nuclear families and each family contains only one affected and one unaffected sibling. Otherwise, it is a valid test of linkage only.
X-Linked Version of Tests
For markers from the X-chromosome that are specified in the XLVAR statement, the preceding tests are not applicable since females have two alleles at such markers and males have only one. Horvath, Laird, and Knapp (2000) present X-linked versions of the TDT, S-TDT, combined S-TDT, and RC-TDT to accommodate these markers. For the X-TDT, the only difference in calculating the values and is that for X-linked markers, transmissions only from heterozygous mothers, instead of heterozygous parents, are used. Note that even though the paternal genotype is not directly used, it must be nonmissing except for when including transmissions to sons in the family, or for daughters who are heterozygous but with a different genotype than their mother (not possible for a biallelic marker).
For the XS-TDT, each sibship is divided into two subsibships so that female sibs and male sibs are analyzed separately. The statistic is then constructed treating the subsibships independently. For female sibs, the parameters  and
and  are the same as those defined for the S-TDT. For male sibs, the X-linked expected value and variance of the number of alleles in affected siblings is calculated across male subsibships as
are the same as those defined for the S-TDT. For male sibs, the X-linked expected value and variance of the number of alleles in affected siblings is calculated across male subsibships as
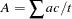 |
and
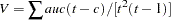 |
where is the number of alleles among all males in a subsibship. The X-linked version of the combined S-TDT is calculated analogously to the combined S-TDT for autosomal markers by using the X-linked versions of the TDT and S-TDT.
The X-linked RC-TDT can be divided into four situations:
Both parents are typed and the X-TDT can be applied.
Only the maternal genotype is missing.
Only the paternal genotype is missing.
Both parental genotypes are missing.
(Note that the first situation also includes the preceding exception when the maternal genotype is nonmissing). Horvath, Laird, and Knapp (2000) show, as with the original RC-TDT, expected values and variances of the number of alleles in affected children when reconstructing parental genotypes in each of the last three situations listed. Using these values, the XRC-TDT can be formed identically to the statistic for the RC-TDT shown in the preceding section.
Permutation Tests
By default, -values from the asymptotic 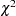 distribution with appropriate degrees of freedom are reported for all tests. However, if the PERMS= option is specified in the PROC FAMILY statement, then Monte Carlo estimates of exact -values are calculated using the permutation procedure for the TDT, S-TDT, SDT, and combined S-TDT and SDT. When the TDT is being performed, including when it is performed in the combined tests, new samples are formed by permuting the alleles that are transmitted to the offspring from the parents and those that are not transmitted (Kaplan, Martin, and Weir 1997). Each affected child in a nuclear family is assigned a genotype comprising one allele from each parent, with each allele being randomly selected from the pair possessed by an individual parent. When the sibling tests are used and the parental information is ignored, the permutation procedure involves randomly permuting the affection status of siblings within each sibship (Spielman and Ewens 1998; Monks, Kaplan, and Weir 1998). For each test, the corresponding test statistic is calculated for the original sample as well as each of the permuted samples. The approximation to the exact -value is then calculated as the number of times the test statistic from a permuted sample exceeds the test statistic from the original sample.
distribution with appropriate degrees of freedom are reported for all tests. However, if the PERMS= option is specified in the PROC FAMILY statement, then Monte Carlo estimates of exact -values are calculated using the permutation procedure for the TDT, S-TDT, SDT, and combined S-TDT and SDT. When the TDT is being performed, including when it is performed in the combined tests, new samples are formed by permuting the alleles that are transmitted to the offspring from the parents and those that are not transmitted (Kaplan, Martin, and Weir 1997). Each affected child in a nuclear family is assigned a genotype comprising one allele from each parent, with each allele being randomly selected from the pair possessed by an individual parent. When the sibling tests are used and the parental information is ignored, the permutation procedure involves randomly permuting the affection status of siblings within each sibship (Spielman and Ewens 1998; Monks, Kaplan, and Weir 1998). For each test, the corresponding test statistic is calculated for the original sample as well as each of the permuted samples. The approximation to the exact -value is then calculated as the number of times the test statistic from a permuted sample exceeds the test statistic from the original sample.
Copyright © 2008 by SAS Institute Inc., Cary, NC, USA. All rights reserved.